力学中的时空变换与DNA弹性杆模型

来源:刘延柱科学网博客,作者:刘延柱。
直梁或曲杆,细长的弹性体在工程技术中并不少见。研究这类弹性体变形规律的传统方法基于17世纪末伯努里 (Bernoulli,J.) 的理论。即建立以变形前梁的轴线为坐标轴的笛卡尔坐标x 和时间t 为自变量,变形后轴线偏移的挠度y(t) 和z(t) 为未知变量的微分方程(图1)。在截面的平面假定和小变形条件下,列写一小段截面的平衡方程,线性化处理后计算挠度的变化规律,截面的转角由挠度对坐标的导数确定。

图1 直杆变形的笛卡尔坐标表达
上述一维弹性体的传统研究方法是材料力学课程的重要内容,也是工程设计的重要理论工具。随着社会的发展,工程实践中有各种大变形的细长弹性体出现,如纤维、缆绳、电缆、钻杆等。这类物体因变形太大,不可能再用传统方法处理。若要另辟蹊径,必须打破对一维弹性体传统概念的思维定势。
暂以圆截面弹性细杆为对象。将杆截面视为刚体,设想此截面沿变形后的杆中心线移动,从杆的起始端到末端,移动过程中截面在空间划过的轨迹与杆的实体互相吻合。沿中心线建立弧坐标s,截面在沿弧坐标s 移动的过程中,相对中心点的姿态不断改变。若将截面在空间中划过的轨迹代替弹性杆的实体,则只要了解刚性截面相对中心点的姿态随弧坐标s 的变化规律,就能确定弹性杆变形后的几何状态(图 2)。
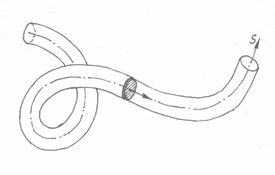
图2 弹性杆的截面沿弧坐标s 的姿态变化
关于弹性杆变形的这一新概念,最早是由德国物理学家基尔霍夫 (Kirchhoff,G..R.) (1859年)和法国物理学家柯塞拉特 (Cosserat,E.)(1909年)提出[1]。由于刚体绕固定点转动问题是早已研究透彻的经典力学问题。因此,只要将刚体定点运动的动力学方程,即欧拉方程的时间自变量t 改为弧坐标s 的空间变量,弹性细杆变形的静力学问题就能利用刚体定点运动的研究成果迎刃而解。基于此思想,1859年基尔霍夫建立了弹性杆大变形问题的 “动力学比拟” 方法(图3)。1927年出版的洛弗 (Love, A.E.H.) 的经典弹性力学著作对此理论有详细的论述[2],但在国内似乎未引起充分注意。

图3 基尔霍夫 (Kirchhoff,G..R. 1824~1887)
关于刚体绕固定点转动问题,1760年欧拉 (Euler,L) 以固定点O 为原点,建立与刚体固结的主轴坐标系 (O-xyz)。将刚体对O 点的动量矩L 表示为刚体对O 点的惯性张量J 与角速度矢量ω 的标量积L=J˖ω,将刚体上作用的力矩M,代入动量矩定理 dL/dt=M。投影至 (O-xyz),设刚体的主惯性矩为A、B、C,得到以角速度ω 的投影ωx、ωy、ωz 为未知变量的微分方程,即欧拉方程:

欧拉方程是奠定经典刚体动力学基础的微分方程。
要将刚体随时间t 的运动转化为沿弧坐标s 进行,相关的运动学概念必须作相应的改变。刚体的角速度定义是无限小角位移ϕ 对时间t 的导数,ω=dϕ/dt。将自变量t 以s 代替,其物理概念必然发生改变。设 (O-xyz) 为圆截面弹性杆刚性截面的主轴坐标系,O 为圆截面的中心,x 和y 轴与截面固定,z 轴沿杆中心轴。依据曲线的微分几何概念,刚性截面绕x 和y 轴的无限小角位移ϕx 和ϕy 对弧坐标s 的导数是曲线沿x 和y 轴的曲率,截面绕z 轴的无限小角位移ϕz 是曲线的扭率。因此,变换后的矢量ω=dϕ/ds 是包含曲率和扭率在内的物理量。文献中未能找到合适的名词,暂称之为弯扭度。
讨论一长度为L 的细长弹性杆,杆的起始端和终止端分别记为P0 和PL。设P 为杆中心线上的任意点,P΄ 为无限接近的邻近点,P 和P΄ 点相对固定参考点O 的矢径分别为r 和r Δr,相对P0 点的弧坐标分别为s 和s Δs。考虑PP΄ 微元弧段内杆的平衡,设P΄ 截面作用的内力主矢和主矩为-F 和-M 。若忽略杆的体力,则P 截面的内力主矢为F,与P΄ 截面的-F 平衡。P 截面的主矩变为M ΔM(图4)。增量ΔM 应满足
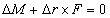
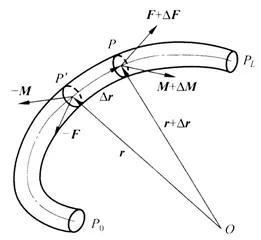
图4 弹性杆微元弧段的平衡
将各项除以Δs,令Δs→0,由于Δr=Δses,es为沿弧坐标的基矢量。导出杆的平衡方程:
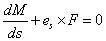
将参考坐标系 (O-xyz) 的原点O 移至P,以 (P-xyz) 为求导的参考坐标系,将上式写作
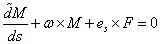
其中,波浪号表示变量相对动坐标系 (P-xyz) 的局部导数,F 为常矢量,ω 为 (P-xyz) 对弧坐标s 的变化率,即上文中定义的弯扭度。
为使沿杆中心线的弧坐标s 不受外力影响,忽略内力F 所引起的拉伸和剪切变形。设杆无原始曲率和扭率,主矩M 与弯扭度ω 之间应满足
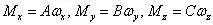
其中,A、B 为截面绕x 轴和y 轴的抗弯刚度,C 为截面绕z 轴的抗扭刚度。等截面杆的刚度系数为常数,取决于杆的弹性常数和截面的几何形状:A=EIx,B=EIy,C=EIz。其中E、G、ν 为杆的杨氏模量、剪切模量和泊松比,杆为均匀各向同性时有G=E/2(1 ν)。对于半径为a 的圆截面杆,Ix=Iy=πa4/4 为截面相对x 轴和y 轴的惯性矩,Iz=πa4/2 为截面相对z 轴的极惯性矩。设α、β、γ 为矢量F 相对 (P-xyz) 各轴的方向余弦,矢量方程
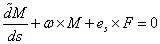
在 (P-xyz) 中的投影式为
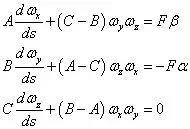
将此方程组与欧拉方程比较,与F 有关的右项视为特殊的力矩分量,二者在形式上完全一致。区别仅在于时间自变量t 被弧坐标s 代替,欧拉方程中的角速度ω 改为弹性杆的弯扭度,其投影ωx、ωy 为杆的曲率,ωz 为杆的扭率。欧拉方程中的主惯性矩A、B 改为表示弹性杆的抗弯刚度,C 表示抗扭刚度。
非线性的欧拉方程在三种特殊情况下存在精确的解析积分,即1760年的欧拉情形。1788年的拉格朗日 (Lagrange,J.L) 情形和1889年的柯瓦列夫斯卡娅 (Kovalevskaya,S.) 情形。依据上述两种方程之间存在的相似性,刚体动力学的所有分析方法和结论均能直接用于弹性杆变形的计算。基尔霍夫动力学模拟方法的基本思想就是通过时空转换,将一维弹性杆的静力学问题转化为刚体动力学问题。对基尔霍夫弹性杆模型的具体计算则依靠对弧坐标s 的数值积分。
基尔霍夫动力学模拟方法的最大优点是刚性截面沿弧坐标的转动角度不受限制,即弹性杆的变形不受限制。即使对弹性细杆反复缠绕折叠的超大变形也能处理不误。因此,能成功地用于缆绳、钻杆、纤维等细杆大变形问题的计算。但基尔霍夫动力学模拟方法从历史文献变为研究热点的推动力,也是最大的用武之地却是似与经典力学无关联的分子生物学领域。
20世纪中期Watson 和Crick 提出了DNA 分子的双螺旋三维结构模型。DNA 是由两条螺旋形戊糖-磷酸骨架和联系骨架的碱基对组成的双螺旋结构的长链分子。虽然对于分子内部结构的研究属于量子力学范畴,但其宏观几何形态及宏观运动的研究可借助经典力学方法。用具有原始扭率的圆截面弹性细杆作为DNA 的宏观力学模型的理论研究得到了实验的肯定(图5)。借助分子生物学的实验手段和统计物理方法,可以测出DNA 弹性杆模型的杨氏模量、泊松比,以及抗弯和抗扭刚度等力学计算所必需的物理常数。在这种宏观力学模型的基础上,研究DNA 的平衡和稳定性规律,经典力学的基本原理和方法得到了充分应用。人体细胞的最大染色体所含DNA 分子的螺旋直径约为2nm,而长度可达7cm,杆长为半径的3.5x107倍。此细长分子链经过反复缠绕、卷曲和折叠,方能被容纳在半径仅10μm 的狭小的细胞核空间内(图6)。因此,DNA 的弹性杆模型以其极端细长性和超大变形,而完全不同于传统弹性力学的研究对象。自上世纪70年代以来,关于DNA 力学模型的研究逐渐形成一支力学与分子生物学交叉的新学科分支,即应用传统的力学分析与分子生物学的实验研究相结合的方法,讨论DNA、RNA以及细菌纤维等各种生物大分子链的几何形态和稳定性问题[3]。
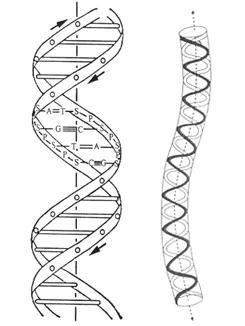
图5 DNA双螺旋结构的弹性杆模型
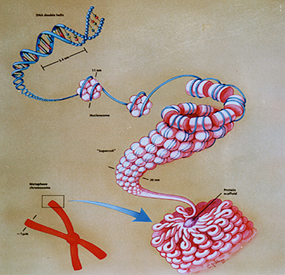
图6 DNA弹性杆模型的超大变形
基尔霍夫动力学模拟方法表明,时空变换扩大了力学问题的适用范围,产生出新的物理概念。与刚体动力学类似,分析力学和运动稳定性等学科中的时间变量也能置换为空间变量,形成“弧坐标分析力学”和“弧坐标运动稳定性”的新概念。
参考文献:
[1] Kirchhoff G. über das Gleichgewicht und die Bewegung eines unendlich dünnen elastischen Stabes. J. Reines Angew. Math., 1859, 56: 285-313
[2] Love A E H. A treatise on mathematical theory of elasticity. 4-th ed., New York: Dover, 1927
[3] Benham G J, An elastic model of the large-scale structure of duples DNA. Biopolymers, 1979,18: 609~623


